Quelle: Entomologie heute 20 (2008): 173 - 191 (Klick für das .pdf)
A Diversity Analysis of Adult Diptera Found on Decomposing Piglets inside and on the Edge of a Western German Woodland inspired by a Courtroom Question*
Wie definiert sich ein Waldrand? Eine durch eine Gerichtsverhandlung angeregte Untersuchung adulter Dipteren auf verwesenden Schweinen im Innern und am Rand eines Waldes im Westen Deutschlands*
ANJA FIEDLER, MARKUS HALBACH, BRADLEY SINCLAIR & MARK BENECKE
* Minor parts of this article were presented as lectures at the meeting of the EAFE (European Association for Forensic Entomology), Lausanne, Switzerland, April 27-29, 2005, and at the 6th ISALM meeting (International Symposium on Advances in Legal Medicine), Hamburg, Germany, Sept. 19-24, 2005. For this publication, the paper was very substantially changed.
Summary: Due to the presence of an adult dead dark-winged fungus gnat (Bradysia sp., Sciaridae), a fly known to live on the borders of forests, on a corpse that was found on the edge of a forest in Western Germany, we were asked during the trial if we could define what the edge of a forest is.
Using two piglets as models, we determined parameters to approach such a definition. The most obvious difference between the inside and the border of the forest was the outside temperature, and the inside temperature of the decomposing piglets, which was higher in the piglet at the border. Over all, 957 individual flies of 56 species of 12 families (Anthomyiidae, Calliphoridae, Drosophilidae, Dryomyzidae, Fanniidae, Heleomyzidae, Lauxaniidae, Muscidae, Piophilidae, Phoridae, Sepsidae, Sphaeroceridae) were found. Specimens of the Drosophilidae and Sepsidae were found exclusively at the edge of the forest and those of the Lauxaniidae, Phoridae, and Sphaeroceridae inside the wood. 36% of the species were found exclusively on the corpse on the border of the forest whilst 10% of the species were exclusively found on the corpse inside of the forest. Several dipteran species showed a clear preference for the border of the forest (Calliphora vicina, Lucilia sericata, Fannia canicularis, Hydrotaea ignava, Liopiophila varipes, Parapiophila vulgaris, Stearibia nigriceps), or for the inside of the woods (Neuroctena analis, Fannia nigra). Succession waves were represented by proportion of insect groups, depending on the location of the piglet. Large numbers of heleomyzids (especially Neoleria sp. 2) were found during the post-decay stage on both corpses. Lucilia ampullacea (Calliphoridae) arrived very early on the habitats, whilst Lucilia caesar arrived 1-2 days later. This is of relevance for post mortem interval analysis. Succession waves were also represented by the relative numbers of adult individuals on the corpses. Here, the relative decrease of calliphorid individuals compared to the increase of heleomyzids, piophilids and dryomyzids was one of the most prominent markers. The decay stages showed a marked difference in the fauna between the two habitats. This was observed not only in the common but also the rare species (Rényis’ diversity). The decomposing piglet on the border was inhabited by a significantly higher number of individuals (576 vs. 381) as well as species (50 vs. 36) of Diptera compared to the piglet inside of the forest. The overall differences in sexes were 23 male/301 female (Calliphoridae), 4 male/89 female (Muscidae), 1 male/87 female (Fanniidae), 116 male/41 female (Piophilidae), and 18 male/238 female (Heleomyzidae). Male heleomyzids and piophilids, in contrast to all other monitored flies, seemed to try to establish a small territory on the corpse, whilst female heleomyzids and piophilids preferred to stay in the sun. They only approach the corpse to lay their eggs, to feed and probably to mate.
Keywords: Forensic Science, Diptera, Heleomyzids, Bradysia, Neoleria
Zusammenfassung: Aufgrund des Fundes einer erwachsenen, toten Pilzmücke mit dunklen Flügeln (Bradysia sp., Sciaridae), die normalerweise an Waldrändern vorkommt, auf einer Leiche am Rand eines Waldes im Westen Deutschlands wurden wir während der Gerichtsverhandlung von der Verteidigung und von der Staatsanwaltschaft gefragt, was als „Waldrand“ angesehen wird. Über zwei Ferkelleichen haben wir versucht, Parameter für eine solche Definition zu erarbeiten. Der offensichtlichste Unterschied zwischen dem Inneren des Waldes und dem Waldrand lag in der Außentemperatur sowie in der Innentemperatur der verwesenden Ferkel, die im Schwein an der Waldgrenze höher war. Insgesamt wurden 957 Fliegen aus 56 verschiedenen Arten und zwölf Familien (Anthomyiidae, Calliphoridae, Drosophilidae, Dryomyzidae, Fanniidae, Heleomyzidae, Lauxaniidae, Muscidae, Piophilidae, Phoridae, Sepsidae, Sphaeroceridae) nachgewiesen.
Die Arten der Drosophilidae und Sepsidae wurden ausschließlich am Rand des Waldes, die der Lauxaniidae, Phoridae, Sphaeroceridae im Wald gefunden. 36 % der Arten fanden sich ausschließlich auf der Leiche am Waldrand, 10 % ausschließlich auf der Leiche im Wald. Mehrere Dipteren-Arten zeigten entweder eine deutliche Vorliebe für den Waldrand (Calliphora vicina, Lucilia sericata, Fannia canicularis, Hydrotaea ignava, Liopiophila varipes, Parapiophila vulgaris, Stearibia nigriceps) oder für das Innere des Waldes (Neuroctena analis, Fannia nigra). Der Anteil der verschiedenen Dipteren-Gruppen in den Besiedlungswellen war von der Lage der Schweineleichen abhängig. Während der Nachverwesungsphase wurden auf beiden Leichen große Mengen von Heleomyziden (vor allem Neoleria sp. 2) gefunden. Lucilia ampullacea (Calliphoridae) tauchte sehr früh auf, während Lucilia caesar 1-2 Tage später erschien. Dieses Verhalten ist von großer Bedeutung für die Bestimmung der Liegezeit.
Nachfolgende Besiedlungswellen waren auch durch die relative Anzahl erwachsener Insekten gekennzeichnet, die sich auf den Schweinekadavern befanden. Ein hervorstechendes Merkmal war vor allem die relative Abnahme von Calliphoriden und die Zunahme von Heleomyziden, Piophiliden und Dryomyziden. Die Dipterenfauna der Verwesungsstufen war auf beiden Schweineleichen deutlich verschieden. Dies wurde nicht nur bei den häufigen, sondern auch bei den seltenen Arten (Diversität nach Rényis) beobachtet. Das verwesende Schwein am Waldrand war von einer höheren Zahl von Individuen (576 im Vergleich zu 381) und Arten (50 im Vergleich zu 36) besiedelt als das Vergleichsschwein im Wald. Das Geschlechterverhältnis war bei den Calliphoridae 23 Männchen/301 Weibchen, bei den Muscidae 4 Männchen/89 Weibchen, bei den Fanniidae 1 Männchen/87 Weibchen, bei den Piophilidae 116 Männchen/41 Weibchen und bei den Heleomyzidae 18 Männchen/238 Weibchen. Im Gegensatz zu allen anderen beobachteten Fliegen schienen die Männchen der Heleomyzidae und Piophilidae zu versuchen auf den Leichen zu siedeln, während die Weibchen es offenbar vorzogen, in der Sonne und im freien Raum zu bleiben. Sie näherten sich den Leichen nur, um ihre Eier zu legen, zu fressen und vielleicht noch um sich zu paaren.
Schlüsselwörter: Kriminalistik, Diptera, Heleomyzidae, Bradysia, Neoleria
1. Introduction
In contrast to fly larvae, which are often used to determine the post mortem interval by estimating the age of the maggots (e.g. MÉGNIN 1894; NUORTEVA 1977; SMITH 1986; LECLERCQ & BRAHY 1990; BYRD & CASTNER 2000; ERZINÇLIOGLU 2000; GOFF 2000; BENECKE 2001a, 2004; GREENBERG & KUNICH 2002), dead adult flies recovered from corpses or other items on crime scenes can be used as indicators of former locations (e.g., deposition sites) of a corpse, and of movement after death.
During a homicide trial in Aachen, Germany, dead adult flies of the genus Bradysia (Sciaridae) had been recovered from a corpse that was found on grassland close to a forest. Bradysia sp. is known to live on the borders of forests (SMITH 1986). Relating to the expert witness statement in the court room (M.B.), defence and prosecution asked how the edge of a forest could be defined. To answer this question ecologically, we monitored all flies during the decomposition of two piglet carcasses inside a forest and on the border of a structurally comparable forest in Western German woodland, approximately 90 km (56 miles) away from the crime scene. For a period of more than 6 weeks (47 days) in autumn 2003, adult flies on the two carcasses habitats were collected to find out if specific fly species or groups were more likely to characterize the border in contrast to the inside of a forest.
2. Material and Methods
2.1. Location
Click on figure above for .pdf
The study was performed from September 16 to November 1, 2003. The experimental area was a forest valley (the Melbtal) south of the city of Bonn in Western Germany (50°42’26”N, 7°05’11” E; elevation 140 m). The climate of Bonn is moderately maritime with an annual average temperature of 11.3 °C, and average annual rainfall of 600 mm. The Melbtal (= valley of Melb) is a mapleash-canyon forest (Aceri-Fraxinetum) composed of Sycamore Maple (Acer pseudoplatanus), Wych Elm (Ulmus glabra), large-leaved linden/lime (Tilia platyphyllos), European Beech (Fagus sylvatica), black alder (Alnus glutinosa), bird cherry (Prunus padus), black elder (Sambucus nigra), common dogwood (Cornus sanguinea), Ranunculaceaen shrubs (Clematis vitalba), Moschatels (Adoxa moschatellina), wood anemones (Anemone nemorosa), Lesser Celandine (Ficaria verna), Herb Paris (Paris quadrifolia), lungwort (Pulmonaria officinalis), Golden Dead Nettles (Lamium galeobdolon), Common Arum (Arum maculatum), horsetail (Equisetum hyemale, E. telamteia), drooping sedge (Carex pendula), European Woodland sedge (Carex sylvatica), wood rush (Luzula sylvatica), woodruff (Galium odoratum), and solomon’s seal (Polygonatum multiflorum) (FRAHM & FISCHER 1998).
2.2. Experimental set-up and determination
Two fresh piglet carcasses (Sus scrofa) were used. One piglet (2.7 kg) was located in an area with a curved shape with a distance of 100 m to the front edge, and 20 m to the side edges of the forest. Nearby was a streamlet. This location was defined as “inner forest” (IF). The second piglet (3.7 kg) was located on the borders of the valley 20 m away from the first location on the border between forest and grass area. This location was defined as “edge of the forest” (EF).
We also recorded the five most obvious decomposition stages (fresh – bloated – decay – post decay – skeletal stage (MÉGNIN 1894; BENECKE 2001B)); and recorded the inner cadaver temperature, and the weight reduction (see below). To avoid loss of maggots, each piglet was put in a plastic tray measuring 80×60 cm. This tray was covered with a dense net to allow evaporation of water. Cages measuring 100×80 cm (grid size 2 cm) kept animal scavengers off. For weighing, the piglets were put on a scale on their plastic trays. Over a period of 47 days, adult insects resting on the corpses were caught with a net (by hand) once a day at random times.
The flies were stored in 70% ethanol and determined to the lowest taxonomic level possible (including determination of sex; here on the family level (see fig. 3 [all figures are visible in the .pdf])) using current determination keys (HALL 1948; LINDER 1949; D´ASSIS FONSECA 1968; MCALPINE 1977; MICHELSEN 1980; DISNEY 1983, 1994; SKIDMORE 1985; BEI-BIENKO 1989; ROGNES 1991; GREGOR et al. 2002; PONT & MEIER 2002) as well as the extensive insect collection of Muse um of Natural History Alexander Koenig in Bonn, Germany.
2.3. Data Logging
The state of decomposition and the feeding behaviour of the flies were documented photographically every day (Nikon Coolpix 700/E2100). Weight reduction of the carcasses was determined by a scale with a precision of 10 g. Data loggers (Eltek 1000 Squirrel) were used for measurement of environmental variables in 10 min intervals with automatic determination of hourly mean values (six Eltek HMP45A & HMP 45D sensors, positioned near the carcasses to detect outside temperature and relative air humidity, and the temperature sensors were placed 25 cm inside of the rectum of the pig to determine inner temperature).
2.4. Ecological indices and statistics
Insect diversity was determined within habitats (α-diversity) and compared between habitats (β-diversity) using the Shannon-Wiener index (H) and the evenness (α-diversity) as well as Renkonen’s index (β-diversity) (WHITTAKER 1967; PIELOU 1969; MAGURRAN 1988). The Shannon-Wiener index (H) is determined by the number and the abundance of each species. If species are homogeneously distributed within the habitat, the index is 1.
The index increases or decreases when species are over- or underrepresented. Renkonen’s percentage similarity index is a measure for relative species abundance and becomes smaller when diversity increases. An additional comparative analysis on dipteran diversity in both habitats was performed using Rényis’ diversity. It compares rare and common species within a parametric scale. To calculate the diversity, we used the software DIVORD 1.50 (PATIL & TAILLIE 1979; TÓTHMÉRÉSZ 1993, 1998; KREBS 1999). Determination of significance (p) was performed by using the Student t-test.
3. Results
3.1. Environmental Influences
3.1.1. Outside temperature
The air temperature during the experimental period was on average 1 °C higher at the edge of the forest (10.5 °C) compared to the temperature inside of the forest (9.6 °C; p<0.001 with Student t-test). Minimum temperatures were observed during the morning hours around 9 a.m., maximum temperatures were reached in the late afternoon around 5 p.m. The highest temperature differences between both habitats existed during periods with the highest absolute temperatures (max.: 24.9 °C (IF) vs. 30.3 °C (EF)). The mean maximum temperature was 13.7 °C (IF) and 16.3 °C (EF). The lowest differences in temperature were detected during periods of lowest temperatures (min.: -1.5 °C (IF) vs. -1.8 °C (EF)). The average minimum temperature was 6.1 °C (IF) and 6.2 °C (EF). The temperature differences between maximum and minimum temperatures showed higher fluctuations at the edge of the forest than inside of the woods (Fig. 1).
3.1.2. Relative humidity
The relative humidity (RH) was on average 5% higher (p<0.001; Student’s t-test) inside of the forest (90%) compared to the edge (85%). Maximum values of humidity were recorded during the morning hours (~ 9 a.m.), minimum values occurred in the late afternoon (~ 5 p.m.). The highest difference in humidity between both habitats was observed between minimum values: the absolute minimum was 52.4% (IF) and 29.2% (EF), the mean minimum value was 75.4% (IF) and 62.4% (EF). Absolute humidity was close to full saturation and did not differ (96.6% IF and 96.9% EF). The differences in air humidity between maximum and minimum values show higher fluctuations at the edge of the forest than inside the wood (Fig. 1).
3.1.3. Inner temperature of the corpses
The mean inner temperature of the piglet (EF) was approximately 2.2 °C higher than the piglet (IF) (EF: 14.1 °C; IF: 11.9 °C). The highest difference in inner temperatures was observed at 5 p.m., the smallest differences were found at 8 a.m. (Fig. 2). The inner temperature of both carcasses increased quite constantly from day 2 to day 4. Over the following three days, the temperature of both corpses started to increase rapidly, even over the outside air temperature. In these days, absolute maxima of the inner temperatures were observed (piglet (EF): 44.6 °C; this was nearly 10 °C higher than the maximum temperature of piglet (IF): 35.7 °C). The difference between the maximum and minimum inner temperature reached highest values during those periods (EF: 11.5 °C; IF: 6.2 °C).
The absolute inner temperature maximum was reached approximately 8 h earlier in the corpse at the edge of the forest compared to the corpse inside of the woods. After day 8, the temperature of both piglets continuously decreased. During the days of the steepest increase in inner temperatures (day 5-7), the air temperature behaved accordingly: the maximum value of inner temperatures was always observed approximately 3 h after a maximum in the air temperature (Fig. 2). Over the whole duration of the experiment, the inner temperatures (EF: 14.1 °C; IF: 11.9 °C) were markedly higher than the air temperatures (EF: 10.5 °C; IF: 9.6 °C). The amplitudes of fluctuations of the inner temperatures were always smaller compared to the fluctuations in air temperature.
3.2. Diptera communities
3.2.1. Visitor spectrum over complete experimental period
Click on figure above for .pdf
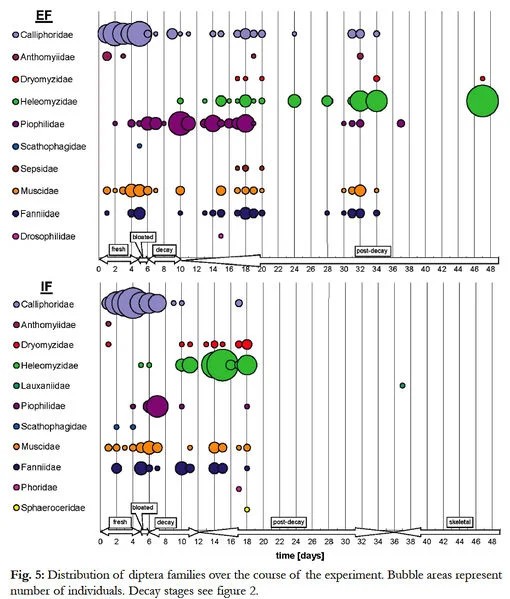
During the experimental period 957 individual flies of 13 families and 56 species were caught on both corpses altogether. 576 individuals (10 families, 50 species) were found on the piglet (EF) and 381 individuals (11 families, 36 species) were found on the piglet (IF). Table 1 [all tables are visible in the .pdf] gives the detailed number of species and individuals of the families in each habitat.
Lauxaniidae, Phoridae, Sphaeroceridae, Drosophilidae and Sepsidae were present even at the edge of the forest or in the inner forest, but were never caught in both habits. A list of species is given in table 2. 36% of the species (from seven families) were exclusively caught on the piglet (EF), including the families Drosophilidae and Sepsidae. 10% of the species (from five families) were only found on the piglet (IF), including the families Lauxaniidae, Phoridae and Sphaeroceridae.
Other families, found on both corpses, are: Calliphoridae, Heleomyzidae, Fanniidae, Muscidae, Anthomyiidae, Piophilidae, Dryomyzidae and Scathophagidae. The most dominant families and species during the complete experiment were: Calliphoridae, which formed the largest group on both corpses (EF: 177, IF: 147). A total of eight species were collected, seven of them could be found on the piglet (EF), but only 5 species were found on the piglet (IF). The most prevalent species was Lucilia caesar with 90 (EF) and 65 (IF) individuals, respectively. Heleomyzidae, which make the second largest number of individuals on both corpses (EF: 144, IF: 112). Four species were present: all four on piglet (EF), two on piglet (IF). Neoleria sp. 2 was the most frequent species in both habitats (EF: 137, IF: 109).
Piophilidae were the third largest group (piglet (EF) with 129 individuals from four species). The most common species amongst them was Parapiophila vulgaris with 73 individuals. For piglet (IF) the third largest group were Fanniidae with 43 individuals. Eight out of nine species of this family were caught at piglet (IF). With 22 individuals, Fannia nigra was the most abundant species. On piglet (EF), Fanniidae were represented by 45 individuals from seven species; Fannia monilis was the most frequently found species (13 individuals).
At piglet (IF) Piophilidae were present with 28 individuals out of four species (15 Liopiophila varipes individuals). Also Muscidae were strongly represented (EF: 59 individuals, IF: 34 individuals). This group also contained the highest overall number of species (19). On piglet (EF), all 19 muscid species were present, whereas only ten species were collected from piglet (IF). The most frequent species was Hydrotaea ignava (13 individuals; edge of the forest), and Hydrotaea similes (9 individuals; inside of the woods). Some Anthomyiidae. (four species, eight individuals) were caught on the edge of the forest (e.g., Egle sp. and Hylemya strenua, three individuals each). Inside of the woods, Dryomyzidae were present (two species; 12 of the 13 individuals were Neuroctena anilis).
3.2.2. Overall distribution of sexes
59% of the flies were female (tab. 2, Fig. 3; Calliphoridae (23 male/301 female), Muscidae (4 male/ 89 female) and Fanniidae (1 male/87 female). Males however dominated in Heleomyzidae (238 male/ 18 female) and Piophilidae (116 male/41 female). Male and female Dryomyzidae were equally distributed (9 male/10 female) on the piglets.
3.3. Decomposition process
3.3.1. Fresh stage
The fresh stage of the two corpses lasted four days. The first eggs were deposited after a few minutes on piglet (EF), and after 30 min on piglet (IF), respectively. On the second day, parts of the piglets were densely packed with eggs. Active larvae were first observed on the third day. The weight of the piglets decreased only slowly until the end of the fresh stage (EF: 98.3%, IF: 96.8%, of original weight).
3.3.2. Bloated stage
The bloated stage lasted until day 5 on both corpses, and was characterized by a high larval activity. The piglet on the edge of the forest was heavily bloated. At this stage the pig let (EF) had a weight of 95%, the piglet (IF) of 97% of its original weight.
3.3.3. Decay stage
The decay stage started at day 6 on and lasted until day 9 (EF), and day 12 (IF), respectively. The piglets were covered by maggots and the heads were nearly skeletonized. Especially piglet (IF) produced a strong decomposition odour. On day 6, the stage with the highest decomposition rate was reached (EF: 1.38 kg/day, IF: 0.54 kg/day). The weight decreased to 57% (EF) and 71% (IF). Over the next days, the progress of the decomposition of the two carcasses changed. More and more maggots left the corpse (EF), which collapsed and started to dry out. At the end of the decay stage, 29% of the original weight had remained. On piglet (IF) the skin was used up and the bones were visible. The consistency of the soft tissue was creamy. Therefore and because maggots dispersed, weight measurements of the piglet (IF) had to be stopped at day 10. At this time, piglet (IF) weighed 37% of its original weight (compared to 21% in piglet (EF)).
3.3.4. Post-decay stage
The post-decay stage of piglet (IF) lasted from day 13 to 36. The soft tissue was increasingly used up and maggots successively migrated away. On day 36, only bones, chondral pieces, hoofs and hairs remained. Post-decay stage of piglet (EF) started on day 10. At the beginning of this stage, the piglet dried out more and more each day and stiffened. Nearly all maggots were leaving the corpse until regular rain from day 15 on resoftened the corpse and made it re-assessable for the maggots. Transition into a fully skeletal stage had not been reached when the experiment finished. From day 47 on, the skin was determined to be mummified. No larval activities of any sort were observed anymore. The mass of the carcass (EF) was 7% at that point.
3.4. Visitor spectrum
Most fly individuals were caught in the postdecay stage (EF: 61%, IF: 34%), followed either by the fresh stage (EF: 26%), or by both the fresh and the decay stage (IF; 29% each) (Fig. 4). Inside of the forest, many species were caught in the decay stage (31%), followed by the fresh and post-decay stage (24% each). The average number of flies caught per day was highest on piglet (EF) during the fresh and the post-decay stage (131/306 individuals, 21/37 species; tab. 3). On piglet (IF) the maximum number of flies per day was caught during the fresh, decay and post-decay stages (110/111/131 individuals, 15/19/15 species) (for details see Figs. 5-7).
3.4.1. Fresh stage
Calliphoridae were the first flies arriving on the corpses; they also represented the largest family during this decay stage (81% EF, 87% IF) (Tab. 4). The most prominent species in the fresh stage were Lucilia caesar (42% EF, 35% IF), Lucilia ampullacea (21% EF, 39% IF) and Calliphora vomitoria (12% EF, 13% IF). An interesting observation was that Lucilia ampullacea arrived very early both on the border and inside of the forest, whilst Lucilia caesar arrived 1-2 days later. Another dominant family in the fresh stage were Muscidae (11% EF, 6% IF), mainly represented by Hydrothaea ignava on the edge of the forest (4%) and Hydrothaea dentipes (3%) inside the forest.
3.4.2. Bloated stage
Like in the fresh stage, the most frequent family on both habitats were calliphorids (74% EF, 76% IF), mostly Calliphora vomitoria (11% EF, 36% IF), Lucilia caesar (54% EF, 32% IF), Lucilia sericata (9% EF) and Lucilia ampullacea (8% IF). Fanniidae were the second most common family on both piglets (EF: 15%, IF: 20%), mostly Fannia canicularis and Fannia immutica (6,5% each) at the edge of the forest, and Fannia monilis and Fannia nigra (8% each) inside the woods. Hydrothaea ignava was the only species of Muscidae on the edge of the forest (11%) (Fig. 4).
3.4.3. Decay stage
On corpse (EF), Piophilids were dominant (66%), mostly represented by Stearibia nigriceps (43%) and Liopiophila varipes (20%), followed by Calliphoridae (33%; four species). Inside of the woods, Calliphoridae were also the most prominent family (30%), followed by Piophilidae (24.5%). Fanniidae (19%) and Heleomyzidae (16%) were only caught inside of the woods.
3.4.4. Post-decay stage
In this stage, the absolute out-standing species was Neoleria sp. 2 (Heleomyzidae) with 40% on the corpse (EF) and 75% on the corpse (IF) (Tab. 4). Parapiophila vulgaris (21%) of the Piophilidae (30%) was another dominant species visiting the corpse (EF). Among other families Fanniidae (9.5%), Calliphoridae (9%) and Muscidae (7%) enriched the Diptera fauna on both piglets. Neuroctena anilis, a Dryomyzidae, makes an additional fraction of 7% on the corpse (IF).
3.5. Diversity analysis
3.5.1. α-Diversity
Over the full course of the experiment, species diversity was higher at the edge of the forest compared to the inside of the forest (H (EF): 4.0/ H (IF): 3.6) even though evenness was similar for both habitats (E: 0.7; tab. 5). The diversity depended on the stage of decay. The lowest diversity was calculated for the bloated stage inside the forest (H: 0.4; E: 0.2). In the fresh and post-decay stage, diversity inside of the forest was lower than that of the edge of the forest. Only in the decay stage, the piglet inside of woods showed a higher Shannon-Wiener index than the edge of the forest.
3.5.2. β-Diversity'
The species assemblage of the carcass on the edge of the forest, and inside of the forest was similar (Renkonen’s percentage similarity index: 70.2%). Relating to the decay stages, the differences in families in both habitats increased with the progression of the decay (Fig. 8). The initial similarity of families (83% in the fresh stage) decreased markedly (55% in the postdecay stage). The fresh stages of the two habitats showed identical values but the other stages were not similar anymore in the end.
3.5.3. Rényis’ diversity
The diversity of the corpse at the edge of the forest was higher not only for the rare (scale parameter is 0), but also for the common species (Fig. 9).
4. Discussion
We found clear faunistic differences on the corpses between both habitats. The diversity strongly depended on the stage of the decomposition. Some species prefer to live on the border between forest and grassland, or inside the woods, and can therefore be used as markers for these habitats. Differences between the sexes of the colonizing flies were also observed and can also be used as markers. The most obvious difference between both habitats was the air temperature (mean ΔT = 1 °C, p<0.001; mean ΔTmax = 2.6 °C). This relates of course to the inside temperature of the two corpses, which was higher at the piglet (EF) (mean ΔT = 2.2 °C; abs. ΔTmax = 10 °C on 7th/8th day of decomposition).
The higher temperatures were due to more direct exposure to sunlight, and they seem to be the main reason for the preference or the avoidance of the habitat by flies. Humidity also differed between the habitats (lower in habitat (EF); mean ΔRH = 5%; mean ΔRHmin = 13%). This was influenced by the outside temperatures: The divergences of the maximum and minimum values of temperature and humidity were always higher at the edge of the forest (Tout: DivabsEF/IF = 32.1/25.8 °C; DivmeanEF/IF = 10.1/7.6 °C; Tin: DivabsEF/IF = 11.5/6.2 °C; RH: DivmeanEF/IF = 67.7/44.2%).
During the experimental period, 957 individual flies of 56 species out of 13 families were found: Anthomyiidae, Calliphoridae, Drosophilidae, Dryomyzidae, Fanniidae, Heleomyzidae, Lauxaniidae, Muscidae, Piophilidae, Phoridae, Sepsidae, Sphaeroceridae. Some of these families were only found at the edge of the forest Drosophilidae, Sepside), or inside of the wood (Lauxaniidae, Phoridae, Sphaeroceridae). 36% of the species were found exclusively on corpse (EF), whilst 10% of the species were exclusively found on the corpse (IF). Several Diptera species showed a clear preference for either the border of the forest (e.g. Calliphora vicina, Lucilia sericata, Fannia canicularis, Hydrotaea ignava, Liopiophila varipes, Parapiophila vulgaris, Stearibia nigriceps), or the inside of the woods (e.g. Neuroctena analis, Fannia nigra). To our knowledge, this topic was not researched yet.
Successional waves were represented by presence and absence, but also by relative amounts of insect groups. These differences depended on the location of the piglets. For example, numerous Heleomyzidae (especially Neoleria sp. 2) were found during the postdecay stage on both corpses. Piophilidae were found mostly during bloated and decay stage inside the forest, but during all stages on the piglet (EF). Additionally it was observed that Lucilia ampullacea (Calliphoridae) arrived very early on the habitats, whilst Lucilia caesar arrived 1-2 days later. This is relevant for post mortem interval analysis.
A much better definition of successional waves arose from the relative number of adult individuals on the corpses. Here, the relative decrease of calliphorid individuals compared to the increase of heleomyzids, piophilids and dryomyzids was one of the most prominent markers. Over the full duration of the experiment, no insect species was alone dominant (α-diversity/ Shannon-Wiener index 3.6 vs. 4; eveness 0.7 vs. 0.7). Both habitats had a similar structure (β-diversity/Renkonen’s percentage similarity index: 70.2%). For the different decay stages, the fauna was however markedly different between the two habitats. This difference was not only caused by the more common, but also by the rare species (Rényis’ diversity).
The decomposing piglet (EF) showed a significant higher number of individuals (576 vs. 381) and species (50 vs. 36) of Diptera compared to piglet (IF). This was not only caused by an excess of an undetermined Neoleria species (Heleomyzidae) at the border of the forest, but was a constant tendency that became the more obvious the further decomposition progressed. In contrast to all other monitored flies, male heleomyzids and piophilids seem to try to establish a small territory on the corpse whilst female heleomyzids and piophilids prefer other sites, mostly sunny ones. They only approach the corpse to lay their eggs, to feed and probably to mate. We are fully aware of the fact that the present study is just a case report containing an analysis that was performed purely out of the necessity to answer a question in an actual homicide trial. Our somewhat rough findings (small piglets, only one experiment) did however help to define what the border of a Western German maple-ash-canyon forest is, and that the presence of Bradysia sp. from the actual case may indeed be restricted to such an ecologically marked border.
We believe that due to the very large number of determined individuals on the determination side, the results did reveal actual ecological properties of the experimental sites, and gave hints for the understanding of the actual homicide case. We also think that our focus on adult instead of larval flies provides an informative yet uncommon approach to a better understanding of the fauna of corpses. Faunistic differences like the ones we found are however most obvious when monitoring adult flies by trapping as well as hand catching and determination which is all very labour intensive and may not always be possible in routine case work.
5. References
Please take a look at the references in the .pdf.